Parede
celular
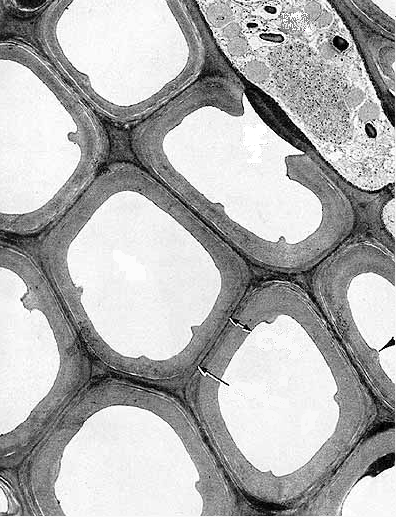
A Célula vegetal elabora uma matriz extracelular complexa que forma um
invólucro que a protege. Foram as paredes das
células do felema da cortiça que foram em 1663 pela primeira vez observadas ao
microscópio, e que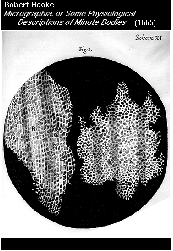 introduziram o próprio conceito de célula. Depois da
referida observação de Robert Hooke muito mais foi apreendido acerca da
estrutura e organização desta estrutura fundamental dos vegetais. Hoje em dia
sabemos que esta estrutura rígida apresenta variações na
paredes que se apõem têm a designação de paredes primárias e são
finas e semi-rígidas. Uma vez terminado o crescimento a parede não necessita
de expandir mais. Nesta circunstância um dos dois casos podem ocorrer, ou se
mantém só a parede primária ou então apõem-se uma parede secundária mais rígida
para o interior da célula. A parede é produzida pela própria célula por
aposição de novas camadas para o interior das mais antigas e a sua
introduziram o próprio conceito de célula. Depois da
referida observação de Robert Hooke muito mais foi apreendido acerca da
estrutura e organização desta estrutura fundamental dos vegetais. Hoje em dia
sabemos que esta estrutura rígida apresenta variações na
paredes que se apõem têm a designação de paredes primárias e são
finas e semi-rígidas. Uma vez terminado o crescimento a parede não necessita
de expandir mais. Nesta circunstância um dos dois casos podem ocorrer, ou se
mantém só a parede primária ou então apõem-se uma parede secundária mais rígida
para o interior da célula. A parede é produzida pela própria célula por
aposição de novas camadas para o interior das mais antigas e a sua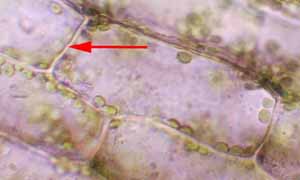 composição
e estrutura é diferente consoante a camada. Para além do papel estrutural ou
“esquelético” da parede celular ela também tem um papel protector, bem
como funções de transporte no interior da célula.
composição
e estrutura é diferente consoante a camada. Para além do papel estrutural ou
“esquelético” da parede celular ela também tem um papel protector, bem
como funções de transporte no interior da célula.
Apesar das paredes primárias
variarem bastante em composição e organização, elas obedecem aos mesmos
princípios gerais: a sua força tênsil deve-se
às longas fibras nelas existentes e
que apresentam enorme resistência compressão devido ao facto dessas fibras se
encontrarem embebidas na matriz extracelular de natureza polissacarídica. As
fibras são constituídas por celulose. A celulose é a macromolécula orgânica
mais abundante da Terra, a restante da matriz é constituída por hemicelulose e
pectina conjuntamente são proteínas estruturais. Todas estas molécula são
mantidas juntas por uma combinação de pontes covalentes que formam uma
estrutura muito complexa.
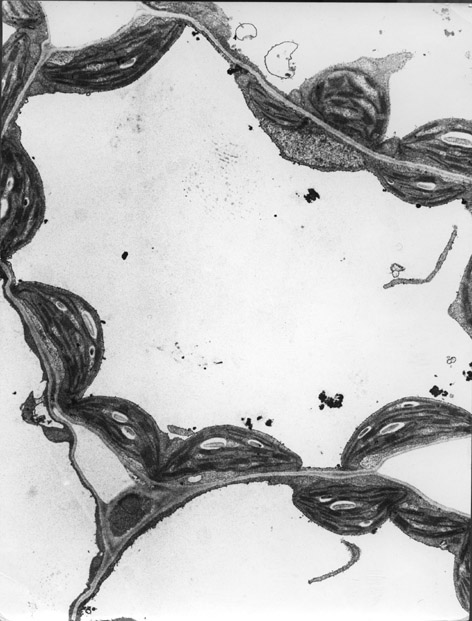
Estrutura
da Celulose
As moléculas celulose são cadeias lineares com pelo menos 500 resíduos
de glicose ligados covalentemente uns aos outros, formando uma fita estabilizada
por pontes de hidrogénio. Para além disto existem pontes de hidrogénio
intermoleculares entre moléculas adjacentes de celulose o que lhes permite
aderir firmemente umas às outras formando feixes de 60 a 70 cadeias de celulose
com a mesma polaridade e organizando-se em agregados cristalinos que tem a
designação de microfibrilas de celulose. Conjuntos de microfibrilas
encontram-se arranjadas em camadas ou lamelas, em que cada microfibrila está
afastada 20 - 40 nm das
microfibrilas vizinhas. Elas estão ligadas umas às outras através de longas
moléculas de hemicelulose que são ligadas a pontes de hidrogénio à superfície
das microfibrilas. A parede celular consiste de várias lamelas arranjadas numa
rede tridimensional.
As hemiceluloses são
um grupo heterogéneo de polissacarídeos ramificados que se ligam à superfície
de cada microfibrila de celulose, bem como a uma outra e por conseguinte ajudam
nas interligações das microfibrilas numa rede complexa. Existem muitas classes
de hemicelulose, mas todas elas têm uma cadeia linear composta por um único
tipo de açúcar a partir da qual se produz pequenas cadeias lineares e outros açucares
quer o esqueleto polissacarídeo quer as cadeias laterais variam de acordo com a
espécie vegetal e o seu estado de desenvolvimento são as moléculas de açúcar
da molécula estrutural que formam pontes de hidrogénio com as microfibrilas da
celulose.
Para além da rede de
microfibrilas celulósicas e das hemiceluloses encontramos uma outra malha
polissacarídea designada por pectinas. Estas são um grupo de polissacarídeos
heterogéneos ramificados que contêm muitas cargas negativas provenientes dos
resíduos de ácido galacturónico. Devido à sua carga negativa as pectinas são
bastante hidratadas e encontram-se acompanhadas de uma nuvem de catiões. Quando
se adiciona o ião cálcio a uma solução de moléculas de pectina, ele forma
pontes entre elas produzindo-se um gel semi-rígido. Certas pectinas são
particularmente abundantes na lamela mediana, região especializada da parede
que cimenta em conjunto as células das paredes adjacentes, e as pontes de ligação
do cálcio são supostas a segurar as componentes da parede celular.
Além
das duas redes dos polissacarídeos, que estão presentes na parede celular do
vegetal, há uma contribuição diversificada das proteínas estruturais.
Biogénese
No final da citocinese, as vesículas golgianas dispõem-se na região
mediana da célula e, após fusão entre si, originam a deposição da matiz.
Estas vesículas são igualmente responsáveis
pelo transporte de material da matriz durante o alongamento da parede
celular primária. Pouco se sabe acerca do seu conteúdo no entanto foi possível
provar a existência de polissacarídeo. A biossíntese da celulose tem lugar à
superfície do plasmalema evolvendo um complexo processo enzimático.